
Behavior of Non-Digestible Polysaccharides in Gastrointestinal Tract: A Mechanistic Review of its Anti-Obesity Effect
- DOI
- 10.2991/efood.k.210310.001How to use a DOI?
- Keywords
- Gastrointestinal tract; obesity; microbiota; non-digestible polysaccharides; digestion; fermentation
- Abstract
Nowadays, obesity in humans is a huge challenge requiring effective, readily available, and low-risk alternative treatments. Intake of edible Non-Digestible Polysaccharides (NDPs) is regarded as a promising strategy to combat obesity. Herein, an overview of the behavior of NDPs in the Gastrointestinal (GI) tract and its association with anti-obesity effect is summarized. Both the in vitro and in vivo of such investigations are discussed. Results show that NDPs can reach the large intestine to mediate the gut microbiota with the extent of regulation varies among NDPs with different chemical structures. The possible anti-obesity mechanisms associated with behavior of NDPs in the GI tract include: (i) inhibition of fat absorption, (ii) modification of gut microbiota and their metabolites such as short-chain fatty acids, (iii) regulation of bile acid profiles, (iv) modulation on appetite and satiety effect, and (v) protection of intestinal barrier. However, more efforts are required to clarify the exact behavior and their beneficial effects of NDPs within GI tract. This paper will provide an overlook of recent advances for better understanding anti-obesity activity of NDPs.
- HIGHLIGHTS
- •
An overview of in vitro and in vivo studies on the behavior of edible Non-Digestible Polysaccharides (NDPs) in the Gastrointestinal (GI) tract is presented.
- •
The anti-obesity mechanisms of NDPs based on their GI behavior are discussed.
- •
- Graphical Abstract

- Copyright
- © 2021 The Authors. Publishing services by Atlantis Press International B.V.
- Open Access
- This is an open access article distributed under the CC BY-NC 4.0 license (http://creativecommons.org/licenses/by-nc/4.0/).
1. INTRODUCTION
Today, obesity, an epidemic with excess or abnormal fat storage, has become a major concern for public health worldwide with rapidly expanding prevalence [1,2]. More than 650 million people are diagnosed with obesity in 2016 according to the World Health Organization [2] and 1 billion people will be influenced by 2030 [3]. Obesity and its multiple complications, such as hyperlipidemia, type 2 diabetes, certain cancers, and so on, all together contribute to reduced life quality, increased mortality, and huge medical expenses [4–6].
An energy imbalance arising from higher energy intake and lower energy expenditure is fundamentally responsible for the occurrence of obesity in addition to other involved factors including genetic metabolism, dysfunctional endocrine, and so on [1,4,6]. To date, strategies proposed to prevent obesity mainly contain anti-obesity drugs, bariatric surgery, diet, and exercise. Among these, the preferred method for fighting against obesity is to persist in regular exercise and a low-energy diet like controlling fat and sugar intake [7]. Nevertheless, the efficiency and success rate of this method has been questioned due to the difficulty of adhering [8]. Besides, anti-obesity drugs previously approved, such as orlistat, sibutramine, and rimonabant, have shown certain adverse effects [4]. Weight loss through gastric bypass or gastric band surgery is more effective than that of drug treatment [8]. However, only a few patients are suitable to adopt this treatment because of its higher risks and cost [4]. Accordingly, effective, readily available, as well as low-risk alternative therapies, have been being explored to treat or alleviate obesity in recent years [8].
As one of the natural products, Non-Digestible Polysaccharides (NDPs) have been in the spotlight of the studies concerning their effects on obesity (Figure 1). They are biomacromolecules composed of monosaccharides linked by glycosidic bonds, and widely distributed in plants, fungi, and microbes [3–9]. NDPs like konjac glucomannans [10], β-glucan [11] have been approved as food supplements for many years. Accumulating evidence has shown that they could prevent the development of obesity through a variety of mechanisms such as regulating lipids metabolism [12,13], reducing inflammation level [14–16], improving insulin resistance [17,18], and so on. The way that NDPs with relatively low bioavailability exert their anti-obesogenic activities in the body, such as changing the body composition, improving disordered metabolism are likely to be based on their behavior in the Gastrointestinal (GI) system to large extent. In this review, the behavior of edible NDPs in the GI tract and their potential anti-obesity mechanisms associated with GI physiology are presented to provide some important insights for better understanding of their control of weight gain.
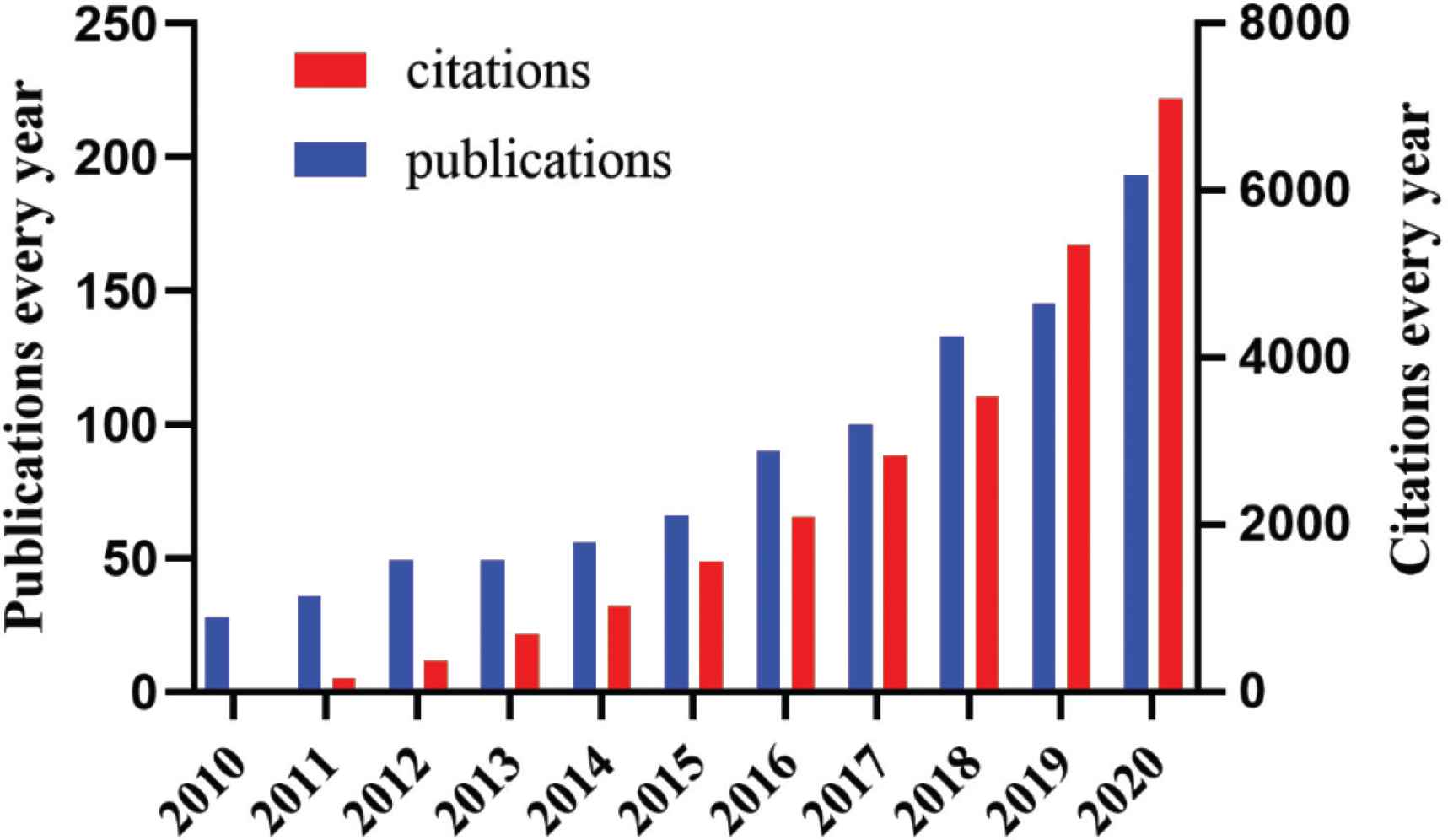
The trend of studies concerning polysaccharides and obesity in the past decade. The data was obtained from the website (http://apps.webofknowledge.com). The theme was set as: “Polysaccharide and obesity”. Retrieve date: February 2021.
2. OVERVIEW OF THE BEHAVIOR OF NDPS IN GI TRACT
In recent years, the digestion and fermentation behavior of natural polysaccharides have been a hot topic for research. Researchers are increasingly interested in if polysaccharides are digestible, how they act on gut microbiota after oral administration, and in what form they are absorbed and functioned [19]. There are many methods proposed to study the changes of polysaccharides and the metabolites produced during the transit in GI tract, including in vitro and in vivo (animals or humans) models.
In terms of the study of digestion, due to the intricacy of our digestive system, in vivo digestive models adopt labeling techniques such as fluorescence labeling [19–21] or digesta collected from animals [22] to investigate the digestibility of polysaccharides. While, in vitro digestion models, either static or dynamic, commonly use digestive enzymes (e.g., salivary, pepsin, gastric lipase, pancreatin), bile salts, and electrolytes. And in vitro digestion models need to control parameters such as pH, temperature, and digestion time to simulate physiological states of the upper GI tract, namely the oral, gastric, and intestinal phase (Figure 2). In contrast, in vivo digestion models are more ideal but are less used for studying digestion than in vitro digestion models owing to the technical difficulties, ethical limitations, and high costs [23]. Besides, there has been a rising interest in the characteristics of their fermentation of NDPs, such as the production of metabolites and alterations of chemical structure, and so on. The fermentation models are divided into in vivo, in vitro static, and in vitro dynamic models. The typical protocols of in vitro fermentation include collecting fresh feces from healthy volunteers without antibiotic treatment, preparing fecal inoculum, and then culturing under an anaerobic environment (Figure 2). Wang et al. [24] have recently reviewed the in vitro static and dynamic fermentation models including the simulator of the TNO in vitro model of the colon (TIM-2) and the human intestinal microbial ecosystem (SHIME®).
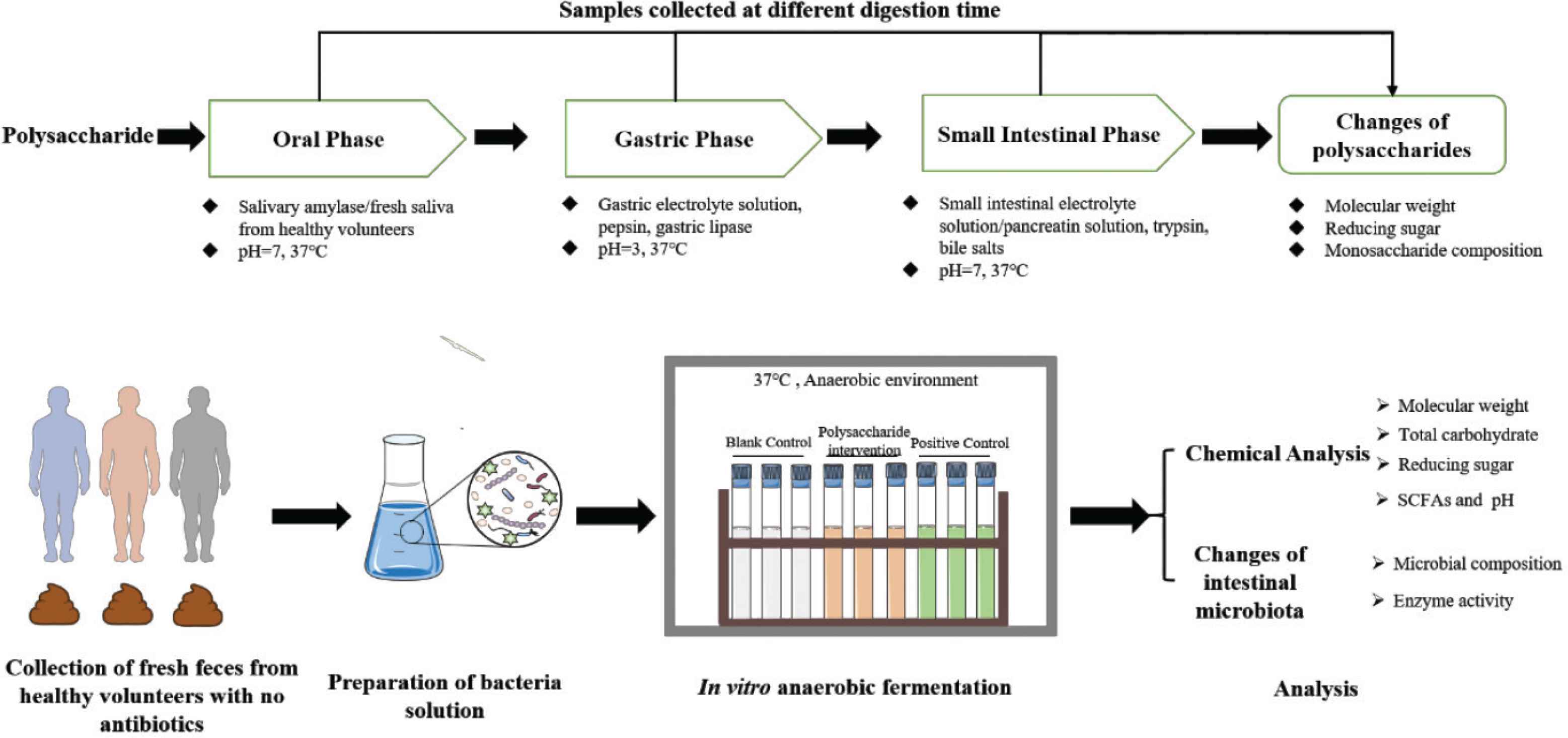
Typical protocols in investigating the digestive behaviors of NDPs in vitro. Images of Erlenmeyer flasks and tubes are referenced from the medical site http://smart.servier.com/ by Servier licensed under a Creative Commons Attribution 3.0 Unported License.
In general, studies of digestive behavior of NDPs focus on changes in molecular weight and reducing sugar contents as well as fermentation by gut microbiota (Table 1). A few studies have also studied the changes in the conformation [25] and viscosity [26] of NDPs in digestion. The experimental results generally varied with each polysaccharide on account of their unique chemical structure resulted from their sources or preparation methods. Some NDPs are partially degraded with a decrease in molecular weight and an increase in reducing sugar, which might be resulted from the effects of pH, enzymes, and bile salts in the process of digestion [27–29]. Reasons for the decline in molecular weight of NDPs within GI tract include the disruption of aggregates and the hydrolysis of glycosidic bonds. However, most of them can resist the digestion of saliva, stomach, and small intestine and can be transited intact into the large intestine [30]. Recently, Li et al. [19] reported that polysaccharide from Dendrobium officinale kept intact during the upper GI tract both in vitro and in vivo, which was similar with many other polysaccharides [31–34]. In the large intestine, since the gut microbiota can generate a variety of carbohydrate-active enzymes [35], these NDPs or their corresponding digestive products (mono-/oligo-/poly-saccharides) can be selectively utilized by gut microbiota to significantly decrease the pH, increase the Short-Chain Fatty Acids (SCFAs), and modify the composition of gut microbiota. Such characteristics of the NDPs match with those criteria of prebiotics [36,37]. Furthermore, the health-promoting activities of polysaccharides are related to their fermentation by microbiota [9,38]. Nonetheless, the exact GI digestion behaviors, particularly the interactions between these polysaccharides or oligosaccharides and intestinal microbiota, still need further exploration. These above studies offer significant information for the diverse applications of polysaccharides in the food industry [23,27,29,30].
| Source | Type | Chemistry and structural information | Study methods | Alterations of molecular weight | Changes in reducing sugars contents | Change of total SCFAs | References | ||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Mouth | Stomach | Small intestine | Mouth | Stomach | Small intestine | ||||||
| Fortunella margarita | Galactoglucan | With a backbone structure of → 4)-β-Glcp-(1 → 4)-α-Glcp-(1 → 4)-α-Galp-(1 → and a side chain of β-Glcp-(1 → 4)-β-Manp-(1 → 6)-β-Glcp | In vitro | — | ↓ | ↓ | N/A | N/A | N/A | N/A | [25] |
| Abelmoschus esculentus | Pectin | Rha, GalA, Gal in a molar ratio of 1:0.76:1.82, Mw = 224.2 kDa | In vitro | ↓ | ↓ | — | — | ↑ | — | ↑ | [24] |
| Artocarpus heterophyllus Lam. (Jackfruit) pulp | N/A | Sugar, proteins, uronic acid in a weight ratio of 79.12:5.83:15.65, mainly composed of Rha, Ara, Gal, Glc, Xyl and GalA, Mw = 1668 kDa | In vitro | ↓ | ↓ | ↓ | ↑ | — | ↑ | ND | [27] |
| Bee collected pollen of Chinese wolfberry | N/A | Man, Rib, Rha, GalA, Glc, Gal, Xyl, Ara in a molar ratio of 0.38:0.09:0.17:0.64:0.22:0.67:0.08:1.03, Mw = 1340.1, 523.2 kDa | In vitro | — | ↓ | — | — | ↑ | ↑ | ↑ | [28] |
| Litchi pulp | N/A | Ara, Gal, Rha and Glc in a molar ratio of 45.2:34.7: 16.7:3.4, with the glycosidic linkages: α- |
In vitro | N/A | ↓ | ↓ | N/A | ↑ | ↑ | ↑ | [29] |
| Mulberry fruit | N/A | Consisted of carbohydrate and uronic acid, Ara, Gal, Glc, Rha in a molar ratio of 35.01:32.12:27.47:3.47, Mw = 128.7, 13.6, 2.9 kDa | In vitro | — | ↓ | — | N/A | ↑ | ↑ | ND | [39] |
| Ganoderma atrum | N/A | Mw = 198.0 kDa | In vitro | N/A | ↓ | ↓ | N/A | N/A | N/A | ↑ | [30] |
| In vivo | N/A | N/A | N/A | N/A | N/A | N/A | ↑ | ||||
| Dendrobium officinale | N/A | Mw = 730 kDa | In vitro | — | — | — | N/A | N/A | N/A | N/A | [19] |
| Gracilaria lemaneiformis | Sulfated polysaccharide | Ara, Fuc, Xyl, Man, Glc and Gal in a molar ratio of 1.14:1.45:7.92:4.23:13.48:71.78, Mw = 22.38 kDa, sulfate (29.82%) | In vitro | — | — | — | — | ↑ | ↑ | ↑ | [31] |
| Fuzhuan brick tea | N/A | Man, Rib, Rha, GlcA, GalA, Glc, Gal, and Ara in a molar ratio of 3.66:1.69:12.11:1.41:28.17:21.97:19.15:11.83, Mw = 828000 g/mol | In vitro | — | — | — | — | — | — | ↑ | [32] |
| Cucurbita moschata | N/A | GalA, Rha, Gal, Ara, and Glc in a molar ratio of 35.93:22.49:21.68:12.05:7.785, Mw: from 12.28 to 270.35 kDa | In vitro | — | — | — | — | — | — | N/A | [40] |
| In vivo | N/A | N/A | N/A | N/A | N/A | N/A | ↑ | ||||
| The flowers of Camellia sinensis | N/A | Man, Rha, Glc, Gal, Xyl, Ara and Fuc in a molar ratio of 2.68:10.17:11.54:22.04:1.49:49.52:2.58 | In vitro | — | — | — | — | — | — | ↑ | [41] |
| Ascophyllum nodosum | Fucoidans | Man, GlcA, Glc, Gal, Xyl, and Fuc at a molar ratio of 16.65:20.34:1.60:9.69:3.44:48.29 | In vitro | — | — | — | N/A | N/A | N/A | ↑ | [42] |
| The seed of Tamarindus indica L. | N/A | Glc, Xyl, Gal, and Ara in a molar ratio of 53.61:25.73:19.79:0.87, Mw = 787 kDa | In vitro | N/A | — | — | N/A | ↑ | ↑ | ↑ | [43] |
| Coralline pilulifera | Galactan | Glc, Gal in a molar ratio of 4.27:95.73, Mw = 716 kDa | In vitro | — | — | — | — | — | — | ↑ | [44] |
| Stichopus variegatus | Fucosylated glycosaminoglycan | Mw = 76.754 kDa | In vitro | — | — | — | — | — | — | N/A | [45] |
N/A, not available, —: no significant difference; ↑ or ↓: increase or decrease significantly before and after each treatment.
Ara, arabinose; Araf, Arabinfuranose; Fuc, fucose; Gal, galactose; Galp, Galactopyranose; GalA, galacturonic acid; Glc, glucose; GlcA, glucuronic acid; Glcp, glucopyranose; Man, mannose; Manp, Mannofuranose; Rha, rhamnose; Rib, ribose; Xyl, xylose.
Digestive behaviors of some natural NDPs
3. ANTI-OBESITY EFFECT VIA INHIBITING FAT ABSORPTION
Obesity happens when energy expended is lower than that consumed. Therefore, it is an effective approach to fight against obesity by controlling the absorption of the energy-dense components in food, especially dietary fat [46]. Dietary fat mainly consists of mixed triglycerides [7]. Starting from the mouth, triglycerides are hydrolyzed, emulsified, and finally absorbed by the small intestine in the form of monoglycerides and free fatty acids [47]. Emerging studies have found that NDPs can significantly affect the process of fat digestion and absorption to prevent obesity via two principal targets, Pancreatic Lipase (PL) and Bile Acids (BAs) micelles.
Pancreatic lipase is a lipase secreted into the small intestine from the pancreas upon food intake [8]. It plays a predominant role in fat absorption in the GI tract, accounting for the hydrolysis of 50–70% of dietary fat [47]. Orlistat, one of the clinically approved anti-obesity drugs, suppresses PL owing to the formation of covalent bonds with the catalytic sites of PL, thus reducing fat absorption [4,8]. But its side effects, such as bloating and diarrhea, can make patients feel uncomfortable [4]. Consequently, developing novel inhibitors from natural products of PL to treat obesity safely has attracted a lot of interest [48,49]. As reported, NDPs possessed inhibitory effects on PL to exert potentially anti-obesogenic effects both in vitro [45,50,51] and in vivo [5,52,53] (Table 2). Remarkably, the inhibitory effects of several NDPs are almost equal to that of orlistat. In a recent study, a barely β-glucan from black Qingke showed strong PL inhibitory effects (IC50 = 2.315 mg/mL) in vitro, compared with orlistat (IC50 = 2.051 mg/mL) [50]. Similarly, the excretion of fecal free fatty acids in High-Fat Diet (HFD) rats was elevated to 21.39–35.86% by black tea polysaccharide extracts compared with orlistat [54]. This indicates that NDPs have the potential to be developed as an effective inhibitor of PL. Although plenty of polysaccharides with inhibitory effect on PL have been found, the precise relationships between structure and activity are not clear now. Currently, there are several probable structural factors concerning the inhibitory activity of polysaccharides on PL. On the one hand, it has been suggested that inhibitory effects on PL by polysaccharides might be attributed to several functional groups in the polysaccharides, including calcium ions [55], free carboxyl groups, sulfate groups, and so on. Zhao et al. [45] recently reported that the inhibitory effect on PL of a glycosaminoglycan was decreased significantly after desulfation or carboxyl reduction, which is similar with the result of desulfation of a galactofucan [56]. As anion groups, the sulfate groups and carboxyl groups might interact with PL via the formation of electrostatic interaction [45]. In addition, it was studied that inhibitory activities of polysaccharides on PL might be positively related to molecular weight and viscosity [50].
| Source | Name | Type | Chemistry and structural information | Methods | Inhibitory effects on lipase | References |
|---|---|---|---|---|---|---|
| Stichopus variegatus | FG | Glycosaminoglycan | With a β1,4-linked trisaccharide repeating unit -{( |
In vitro | Reducing PL activity by 55.4% at 1.25 mg/mL | [45] |
| dsFG | The desulfated FG (the degree of de-sulfation of dsFG > 91%) | In vitro | Lower inhibitory activity on PL compared with FG | |||
| crFG | The carboxyl-reduced FG (the carboxyl groups were reduced completely) | In vitro | Lower inhibitory activity on PL compared with FG | |||
| Laminaria japonica | CGF-3 | Galactofucan | Fuc, Xyl, Gal, Glc, GalA, Man in a molar ratio of 14.9:1.0:16.8:1.7:3.9:6.3, Mw = 527.3 kDa, sulfate groups (26.7%) | In vivo | Decreasing PL activity to 30.70% at 2.00 mg/mL | [56] |
| DSCGF-3 | The desulfated CGF-3 [sulfate groups (6.4%)] | In vitro | Lower inhibitory activity on PL compared with CGF-3 | |||
| Ganyucang | BTHB | β-Glucan | Gal, Xyl, Ara, and Glc in a molar ratio of 0.42:0.81:1.0:27.88, Mw = 331.8 kDa | In vitro | IC50 = 2.315 mg/mL | [50] |
| Dingqing | BLTHB | Gal, Xyl, Ara, and Glc in a molar ratio of 0.25:0.74:1.0:24.05, Mw = 196.3 kDa | In vitro | IC50 = 3.712 mg/mL | ||
| Zangqing 320 | WTHB | Gal, Xyl, Ara, and Glc in a molar ratio of 0.26:0.65:1.0:34.08, Mw = 126.1 kDa | In vitro | IC50 = 7.302 mg/mL | ||
| Codium fragile | CFSP | Sulfated polysaccharide | Gal, Glc, Xyl in a molar ratio of 79.82:18.24:1.94 | In vitro | IC50 = 101.81 μg/mL | [53] |
| In vivo | Reducing the lipase activity in the intestine by 38% after administration at 150 mg/kg of BW daily | |||||
| Cymodocea nodosa | CNSP | Sulfated polysaccharide | Sugar, sulphate, proteins, lipids in a weight ratio of 54.90:23.17:10.50:2.13 | In vitro | IC50 = 108.88 μg/mL | [52] |
| In vivo | Inhibiting the lipase activity by 30% in the intestine after intervention with 200 mg/kg of BW CNSP | |||||
| Cystoseira crinita | CCSP | Sulfated polysaccharide | Ara, Man, All, Gal, Glc, GlcN, Alt, Arap, Araf, Manf, Xylp, Galp, Galf, Ribf in an area ratio of 19:6.9:7.5:8.98:0.4:1.02:7.1:0.5:0.5:1.8:1.6:6:0.3:0.08 by GC–MS analysis | In vitro | IC50 = 96.73 μg/mL | [5] |
| In vivo | Decreasing the intestinal lipase activity by 38% after oral intake at 200 mg/kg of BW daily |
All, Allose; Alt, Altrose; Arap, Arabinopyranose; BLTHB, blue Qingke (Dingqing) β-glucan; BTHB, black Qingke (Ganyucang) β-glucan; CCSP, Cystoseira crinita sulfated polysaccharide; CFSP, sulphated polysaccharide from Codium fragile; CGF-3, a molecular-weight homogeneous fraction of polysaccharide from Laminaria japonica; CNSP, Cymodocea nodosa sulphated polysaccharide; crFG, carboxyl-reduced FG; DSCGF-3, desulfated CGF-3; dsFG, desulfated FG; FG, fucosylated glycosaminoglycan from sea cucumber; Galf, Galactofuranose; GalNAc, N-acetyl-galactosamine; GC-MS, gas chromatography–mass spectrometry; GlcN, glucosamine; Ribf, ribofuranose; WTHB, white Qingke (Zangqing 320) β-glucan; Xylp, xylopyranose.
Inhibitory effects of lipase by NDPs
As a group of amphipathic steroid molecules synthesized from cholesterol by the liver, BAs are secreted into the small intestine to promote the digestion and absorption of foods including fat and cholesterol [8]. Previous studies have shown that polysaccharides binding with BAs could disrupt micelles where BAs act as emulsifiers, affecting the ability of micelle to solubilize the cholesterol, monoglycerides, and free fatty acids [55,57,58]. Subsequently, the absorption of fats and cholesterol within the small intestine can be reduced by polysaccharides, thus decreasing the risk of obesity and its complications [59]. However, despite the interest in the interactions between polysaccharides and BAs or BAs micelles, their nature remains unsure because of several restrictions such as in vivo intricate heterogeneity and limited access to samples, as well as the difficulty in simulating in vivo conditions in vitro [59]. To develop novel natural cholesterol-lowering agents, more efforts should be taken to uncover the mechanisms of actions of polysaccharides on BAs micelles in future studies.
4. ANTI-OBESITY EFFECT VIA FERMENTATION IN THE INTESTINE
As one of the most important prebiotics, NDPs may manipulate gut microbiota and their metabolites, such as SCFAs and secondary BAs, through their fermentation in the intestine to combat obesity and linked disorders (Table 3).
| Source | Type | Chemistry and structural information | Models | Modification of dysfunctional microbiota | SCFAs | Other alterations |
|---|---|---|---|---|---|---|
| Pearsonothuria graeffei [78] | Fucoidan | With 4-O-mono- and 2,4-di-O-sulfated groups and a tetrasaccharide repeating unit | HFD-mice | →alpha diversity, ↓F/B | N/A | Attenuated hyperlipidemia and hyperglycemia |
| ↑Parabacteroides, Alloprevotella, and Olsenella | Protected the liver against steatosis | |||||
| ↓Acetatifactor, Desulfovibrio, Dorea | Reduced serum inflammatory cytokines | |||||
| Decreased macrophages infiltrating into adipose tissue | ||||||
| Orafti, Oreye, Belgium [95] | Inulin-type fructans | Synergy 1, namely, inulin/oligofructose 50/50 mix | Obese women | ↑F/B | N/A | Reduced LPS levels |
| ↑Bifidobacterium, Faecalibacterium prausnitzii | ||||||
| ↓Bacteroides intestinalis, Bacteroides vulgatus, and Propionibacterium | ||||||
| Mytilus coruscus [73] | α- |
With the main chain consisting of (1→4)-α- |
HFD-mice | ↑abundance and diversity, ↓F/B | ↑SCFAs | Protected from non-alcoholic fatty liver |
| ↑Mailhella, Alloprevotella, Butyricimonas, Parabacteroides, Akkermansia, Bifidobacterium | ↑butyrate (in cecal samples) | Suppressed inflammatory factors secretion | ||||
| ↓Allobaculum, Pseudomonas, Hydrogenophaga, Romboutsia, Turicibacter, Ruthenibacterium, Faecalibaculum | Inhibited PPAR-γ and sterol regulatory element-binding protein-1c expression | |||||
| Laminaria japonica [65] | N/A | N/A | HFD-mice | ↑alpha diversity, ↓F/B | ↑total SCFAs (fecal and colonic content) | Decreased the weight of epididymal adipose tissues |
| ↑Rikenellaceae and Bacteroidales S24_7 group | Reduced lipid deposition in adipocytes and hepatocytes | |||||
| ↑Bacteroides spp. and Ruminococcaceae spp. | ||||||
| ↓Pseudomonas spp. and Lachnoclostridium spp. | ||||||
| Kefir grains by the KU Center for Food Safety [72] | N/A | Total carbohydrate (84.1%) | HFD-mice | →alpha diversity, ↓F/B | N/A | Lowered the weight gain and epididymal adipose tissue weight |
| ↑Akkermansia, ↓Adlercreutzia, Coprococcus, Oscillospira, Ruminococcus | ||||||
| Polygonatum odoratum [74] | N/A | N/A | HFD-fed rats | ↑species richness, →F/B | →total SCFAs | Attenuated weight gain, fat accumulation, epididymal adipocyte size |
| ↓Clostridium, Enterococcus, Coprobacillus, Lactococcus, and Sutterella | ↑isobutyric acid, butyric acid, valeric acid (in feces) | Promoted the expression of genes related to adipocyte differentiation and lipolysis | ||||
| Reduced the expression involved in lipogenesis | ||||||
| Baker’s yeast [96] | Glucan | A linear β-(1→3)-glucan: carbohydrate (99%), Mw = 25 kDa | HFD-mice | ↑Bray Curtis diversity, ↓F/B | N/A | Hypoglycemic effects by inhibiting the expression of sodium-glucose transporter-1 |
| ↑Akkermansia | Enhanced glycogen synthesis but inhibited the hepatic fat accumulation | |||||
| Gracilaria lemaneiformis [71] | N/A | N/A | HFD-mice | →alpha diversity, ↓F/B | ↑total SCFAs in the fecal | Decreased weight gain and adipose tissues |
| ↓Enterococcus spp., Pseudomonas spp., Ruminiclostridium spp. | Lowered fat accumulation of adipose tissues | |||||
| ↑Bacteroides spp., Allobaculum spp., Rikenellaceae spp. | ||||||
| Undaria pinnatifida [97] | Sulfated polysaccharides | Man, Rha, GalA, Glc, Gal, Xyl and Fuc in a molar ratio of 22.79:6.13:1.12:8.23:29.85:5.11:26.77, sulfate content (21.59%) | HFD-mice | →species richness, ↓F/B | ↑Acetate, propionate, and butyrate | Improved gut barrier integrity and inflammation |
| ↓Lachnospiraceae_NK4A136 | Decreased intestinal lipid absorption | |||||
| ↑Prevotellaceae_UCG-001 group | Increased intestinal carbohydrate utilization |
→, no significant difference; ↑ or ↓, increase or decrease significantly before and after each treatment;
PPAR-γ, peroxisome proliferator-activated receptor γ; N/A, not available; BW, body weight.
Anti-obesity mechanisms in relation to the fermentation of NDPs in the large bowel
4.1. Modification of Microbiota
Over the past decade, the intestinal microbiota, a vital “microbial organ” composed of trillions of microbes [9], has caught widespread attention with the rapid development of associated sequencing technologies and omics technologies [60,61]. The casual relationship between obesity and intestinal microbiota has been discussed in many studies, among which a piece of evidence strongly shows that the intestinal microbiota from twins with different phenotype concerning obesity induces respective phenotypes in mice [62]. Recently, the intestinal microbiota is generally considered an environmental factor correlating the onset of obesity, although the underlying mechanisms are still not fully delineated [1,63]. NDPs can combat obesity, specifically to enhance the diversity and richness of the microbiota, reduce the ratio between Firmicutes and Bacteroidetes (F/B), and alter specific genus- or species-level.
Mounting studies have found that the fermentation of several polysaccharides significantly increased microbial diversity and richness [64,65]. The diversity of intestinal microbiota in obese individuals has been reported to be lower than that of lean individuals [66]. There is evidence suggested that low richness of intestinal microbiota might be associated with reduced stability of the microbiota, resulted in elevated risk of obesity and its related metabolic disorders [67]. Thereby, a NDP-induced increase of microbial diversity or richness may be of great benefit to improve the dysfunctional microecology in the obese subjects.
In addition, an increase F/B correlates with enhanced capacity for energy absorption which ultimately resulted in obesity [68]. Increased F/B in rodents [69] and humans [70] with obesity was often observed, yet this variation can also be reversed by fermentation of many [71–73], but not all, polysaccharides [74,75]. Apart from these modifications, NDPs also lead to a decrease or increase in bacteria that are positively or negatively associated with obesity respectively through their fermentation in the intestine. For instance, NDP intervention by fucoidan could increase the beneficial bacterium Akkermansia muciniphila [76], which could ameliorate metabolic disorders in mice partly through up-regulating the intestinal levels of endocannabinoids in dietary obese mice [77]. Previous studies also observed some polysaccharides improved obesity along with an increased population of benign microbes such as Alloprevotella spp. [78], Prevotella spp. [79], Parabacteroides goldsteinii [80]. Likewise, it was demonstrated that a water extract of Ganoderma lucidum mycelium (WEGL) could prevent obesity via modifying the gut microbiota, therein including the reduction of Oscillibacter spp., Mucispirillum spp., and Eubacterium fergusonii [81]. These bacteria are in a positive association with inflammation and the development of obesity. And the high molecular weight polysaccharides (>300 kDa) of WEGL was responsible for the anti-obesity activity of the extract [81].
Therefore, polysaccharides-induced alterations of gut microbiota might be of great significance to improve the dysfunctional intestinal microecology in the obese. However, it is noteworthy that the impacts on the gut microbiota of polysaccharides mentioned above might not be all present in a specific study. Meanwhile, further investigations should be paid to clarify the relationship between these impacts and anti-obesity activity by free-germ mice or fecal microbiota transplant studies [80,81] to lay a foundation for targeting gut microbiota to treat obesity and related metabolic diseases using NDPs.
4.2. SCFAs Production
Anti-obesogenic activities of NDPs might be largely due to SCFAs generated during their fermentation. The SCFAs refer to a group of organic fatty acids with 2–6 carbon atoms including mainly acetate, propionate, and butyrate [82], which are involved in regulating physiological activities in local tissues. Firstly, enhanced SCFAs arising from NDP intake will directly lower the luminal pH values, suppressing the growth of pathogens [83]. Besides, SCFAs, particularly butyrate, have the capacity of supplying energy for the intestinal epithelial cells [9,84], and participating in their regulation of proliferation and differentiation depending on concentration [9]. Several studies have elaborated on a protective role of SCFAs in maintaining the intestinal integrity [85–87], which is of great significance for health [88].
The local intestinal effects of SCFAs can determine which signals are subsequently triggered. For example, SCFAs activation of G-protein-coupled receptors GPR41 and GPR43 enhances the intestinal hormones Glucagon-like Peptide-1 (GLP-1) and Peptide YY (PYY) [89]. While these hormones locally regulate the motility of the gut [37], they subsequently enter circulation where they affect other organs to produce satiety, alleviate insulin resistance and inflammation in obese subjects [8].
Short-chain fatty acids can also affect distant organs/tissues (e.g., brain, liver, adipose tissue, and muscle) to exert their metabolic regulatory effects. Among them, adipose tissue is a major target. For example, SCFAs activated the GPR43 receptor in adipose tissue and inhibited the insulin signaling, which suppressed fat accumulation and promoted energy expenditure [90]. Also, acetate and propionate can be metabolized in the liver. Acetate serves as a lipogenic compound while propionate is one of the substrates for gluconeogenesis and can inhibit lipogenesis [83]. More recently, there are a lot of discussions concerning the effects of NDPs on the brain–gut axis [17], in which SCFAs might act as important mediators in interactions between the gut and brain. For example, acetate from colonic fermentation of NDP could cross the blood–brain barrier to directly decline appetite through a hypothalamic mechanism [91]. Above all, SCFAs not only influence the local tissues but serve as key metabolic signals to act on other tissues through a variety of pathways, thereby profoundly regulating metabolic activities. Increased SCFAs might be a key mediator in the impacts of fermentable NDPs.
In conclusion, the fermentation of NDPs in the intestine can be considered as an important target for control of weight gain, which is an outcome of a combination of multiple structural characteristics. NDPs with different molecular weight have different fermentation features. It was suggested that NDPs were more easily by intestinal microbiota when degraded or with appropriate molecular weight [92,93]. And the fermentation of pectin oligosaccharides was reported to be positively related to the branching degree and the content of neutral sugar [94]. Further works in structure–activity relationship should be paid more attention.
5. ANTI-OBESITY VIA REGULATING APPETITE AND SATIETY
Polysaccharides consumption can regulate appetite and satiety to manage food and energy intake, thereby preventing the development of obesity [98,99]. On the one hand, these effects depend in part on their physical effects like the bulking and viscosity effect as well as texture property [100]. The bulking effect does not only result in reduced food energy density and palatability but also contributes to satiety by inducing gastric distension [100]. Besides, some NDPs for example Konjac glucomannans [101] and increase the viscosity of digesta and slow the absorption of nutrients (especially fat which is discussed earlier), thus contributing to delayed gastric emptying, slowed bowel transit time, increased satiety, and reducing further food intake [100,102].
On the other hand, food ingestion induces satiety in the GI tract through mechanical and humoral stimulation. NDPs can regulate the secretion of GI hormones such as GLP-1, PYY, Cholecystokinin (CCK) and ghrelin to control the uptake of food. A recent study found that a NDP from flaxseed contributed to satiety through up-regulation of GLP-1, which might further increase the level of adiponectin and promote lipid metabolism via AMP-Activated Protein Kinase (AMPK) regulation [13]. Several studies also have demonstrated that NDPs intake is positively associated with the level of PYY [103,104] and CCK [105]. CCK can inhibit the activity of hunger-stimulating neuropeptide Y in the hypothalamus [8]. However, some NDPs can decrease the expression of ghrelin that induces hunger via acting on hypothalamic brain cells in the central nervous system [106]. Fermentable NDPs produce SCFAs that are importantly involved in the regulation and control of appetite and satiety through stimulation of hormonal and neural signals in various organs and tissues [107].
6. ANTI-OBESITY VIA REGULATING BA PROFILES
Bile acids, consisting of primary and secondary BAs, play a pivotal role in the regulation of glycolipid metabolism in addition to assisting fats digestion and absorption. They are a group of vital signaling molecules with the ability to modulate energy metabolism by interacting with receptors, Takeda G protein Receptor 5 (TGR5), and Farnesoid X Receptor (FXR) [108,109]. Intestinal TGR5 could be activated by the secondary BAs, which induced GLP-1 secretion in intestinal L cells, alleviated glucose homeostasis, and combated obesity in obese mice [108]. And the inhibition of intestinal FXR could lower serum ceramides and promote beige fat thermogenesis, subsequently alleviating metabolic syndrome [109]. Moreover, the affinity and activation or inhibition potency on BA receptors vary with individual BAs [110]. As a result, BA profiles count to the energy regulation in the host and altered BA profiles have been investigated as a potential anti-obesity mechanism for NDPs.
Importantly, the BA profiles are closely linked to the intestinal microbiota. Bile Salt Hydrolase (BSH) enzymes secreted by some bacteria deconjugate conjugated primary BAs, thus generating unconjugated primary BAs. Unconjugated primary BAs escape uptake into the colon where they are dehydroxylated at 7α position to produce secondary BAs (deoxycholic and lithocholic acid) [110,111]. Other microbial modifications contain esterification, desulfation and so on [110]. Moreover, BSH activity was found to have significant effects on lipid metabolism, weight gain, and cholesterol levels in the host [111]. Gut microbiota dysbiosis and the decreased bacteria with BSH enzymes can result in perturbations of BA profiles and induce diseases related to glycolipid metabolism [112]. NDPs can target intestinal microbiota to regulate the BA profiles in the host. Chen et al. [113] observed that fucoidan could reverse the decreased BSH enzyme activity by modulating gut microbiota in HFD rats, modulate the BA composition, and finally improve the diet-induced dyslipidemia. Another study reported that intervention of a sulfated NDP could decrease the hydrophobic BAs but enhance the hydrophilic BAs might through up-regulating the relative abundance of Prevotellaceae_UCG-001, Corprococcus_1, and Alistipes and Roseburia and Lachnospiraceae_NK4A136_group, respectively [114].
Furthermore, many polysaccharides can bind with BAs, which can suppress the enterohepatic circulation of BAs and affect BA pool size [58,115,116]. Specifically, the administration of a NDP from Ophiopogon japonicus (MDG-1) significantly decreased the serum total BAs levels as compared with the HFD group. Further detection of individual BAs in the urine, liver, and fecal suggested that MDG-1 could absorb BAs in the gut lumen, block the enterohepatic circulation and subsequently decrease the BA pool size in the liver. This inhibited the expression of FXR while activated the transcription of genes CYP7A1 and CYP8B1 involved in the synthesis of BAs in the liver, thus improving hyperlipidemia [117]. Meanwhile, polysaccharides with the capacity to bind with BAs likely provide a larger platform for BAs to interact with microbiota. Hence, the changes in gut microbiota resulting from a supplement of NDP significantly contribute to the BA profiles and metabolism of the host. Based on available evidence, we suggest a polysaccharide/gut microbiota/BAs/FXR or TGR5 axis as a potential mechanism to understand anti-obesity activities of NDPs which still needs further research.
7. ANTI-OBESITY VIA TARGETING INTESTINAL BARRIER FUNCTION
An increase in serum Lipopolysaccharides (LPS) levels has been observed in subjects with diet-induced or genetic obesity [95,118,119], which is called metabolic endotoxemia. It is a chronic condition where elevated LPS in serum can be recognized by Toll-like Receptor 4 (TLR4) on various tissues, subsequently bringing about systemic inflammation and insulin resistance [83,119,120]. There may be many potential causes of metabolic endotoxemia, one of which is that the disrupted intestinal barrier in the obese may allow the intestinal microbes and LPS to enter the circulation [121]. As shown in Table 4, polysaccharides in the intestine can improve the intestinal barrier to alleviate metabolic endotoxemia, hence exerting their anti-obesogenic effects.
| Source | Type | Chemistry and structural information | Models | Daily intake and period | Impacts on the intestinal barrier | Endotoxin | References |
|---|---|---|---|---|---|---|---|
| Fuji apple | N/A | Rha, GalA, Glc, Gal and Ara in a molar ratio of 1.00:17.67:8.50:4.01:2.03, Mw = 2102.472, 135.849, 9.622 kDa | HFD-rats | 5%, 10% (w/v) 10 ml/kg of BW for 14 weeks | Decreased gut permeability with elevated expression of occludin | ↓LBP | [15] |
| Regulating the function of goblet cells through inhibition of Relmβ and autophagy induction | |||||||
| Finger millet CO (Ra) 14 | Arabinoxylan | Rha, Xyl, Man, Gal, Glc in a molar ratio of 35.3:44.7:3.7:9.5:6.8 | HFD-mice | 1.0 g/kg of BW on every alternate day for 10 weeks | Improved colonic barrier function with up-regulation of ZO-1, claudin 2 and 4, Muc2 | ↓LPS | [127] |
| Bamboo-shaving | Arabinoxylan | With a main chain of β-1,4- |
HFD-mice | 400 mg/kg of BW for 8 weeks | Improved intestinal barrier integrity as shown by the complete structure as well as clear and inerratic tight junction under the transmission electron microscopy | ↓LPS | [64] |
| Hirsutella sinensis | N/A | A higher content of Man, Gal, N-GalN, N-GlcN, Rha and Fuc | HFD-mice | 20 mg/kg of BW for 12 weeks | Reduced intestinal permeability in HFD-fed mice along with enhanced expression ZO-1 of the colon | ↓LPS | [80] |
| Poria cocos | β-glucan | 1,3-β- |
ob/ob mice | 1 g/kg of BW for 4 weeks | Maintained intestinal integrity by up-regulating the expression of Muc5, ZO-1, and occludin in the ileum | ↓LPS | [118] |
| Polygonatum kingianum | N/A | Mw = 178.6 kDa, consists of Man, GalA, Gal, and Fuc | HFD-rats | 120 mg/kg of BW for 14 weeks | Reversed the intestinal barrier injuries by elevating ZO-1 and occludin expression | ↓LPS | [131] |
| Apple | Pectin | N/A | HFD-rats | Free access to a standard chow diet with 5% pectin (w/w) for 6 weeks | Maintained gut barrier function by improving the level of claudin 1 | ↓LPS | [132] |
| Enhanced the expression of intestinal alkaline phosphatase | |||||||
| Raphanus sativus | N/A | Rha, Fuc, Ara, Xyl, Man, Gal, Glc, GlcA and GalA in molar ratios of 5.4:5.0:22.9:0.4, Mw = 61 kDa | HFD-mice | HFD with a daily oral gavage of 2 mg/kg for 6 weeks | Improved gut barrier function by improving the level of ZO-1 | ↓LPS | [133] |
| Restored colon shortening |
BW, body weight; w, weight; v, volume; LBP, LPS-binding protein; ZO-1, zonula occludens-1; Muc5, mucin 5.
NDPs that improve intestinal barrier and alleviate metabolic endotoxemia
An intact intestinal barrier is pivotal in separating the gut lumen from the inside of the body besides nutrition absorption. One of the key barriers is the intestinal mucous layer between enterocytes and the intestinal lumen. Generally, increasing the number of goblet cells and their expression of Mucin 2 (Muc2) is the target of certain polysaccharides to strengthen the mucous layer [122–124]. However, a recent study found that a NDP from apple counteracted the HFD-induced goblet cell hyperplasia and reduced thickness of the mucous layer by down-regulating Resistin-like Molecule β (Relmβ) and promoting autophagic process rather than up-regulating Muc2 expression [15].
Additionally, the tight junctions between two neighboring enterocytes act as a vital mechanical barrier closely linked with gut permeability and the translocation of toxins such as LPS [125]. They are composed of transmembrane tetraspan proteins (e.g., occludin, claudins), junctional adhesion molecules, and intracellular scaffold proteins (e.g., zonula occludens-1) [126]. It has been shown that obesity is characterized by increased permeability along with the decreased expression of tight junctions [80,127]. NDPs might promote tight junction related gene expression involving several specific pathways such as inhibition of nuclear factor kappa B/TLR4 [128] and up-regulation of AMPK [129], thus restoring the intestinal barrier and alleviating metabolic endotoxemia in obesity (Table 4). Moreover, as discussed earlier, the fermentation of certain NDPs in the gut lumen could regulate gut microbiota and yield of SCFAs, which may mediate the effect of polysaccharide on intestinal barrier since the alteration in commensal microbiota was reported to be related to metabolic endotoxemia and obesity [120,121,130] and SCFAs can regulate intestinal integrity as mentioned earlier.
8. CONCLUSION AND PERSPECTIVE
In this review, we summarized the behavior of NDPs in the GI tract and highlighted their anti-obesity mechanisms in association with their physiological effects. Results show that NDPs can reach the colon, where they are utilized by colonic microbes. They show the therapeutic effects on obesity within the GI tract via mechanisms including controlling fat absorption, fermentation in the intestine, regulating BA profiles, appetite and satiety as well as improving metabolic endotoxemia by restoring the intestinal barrier (shown in Figure 3). Furthermore, these mechanisms are closely related to other metabolic impacts in the body.
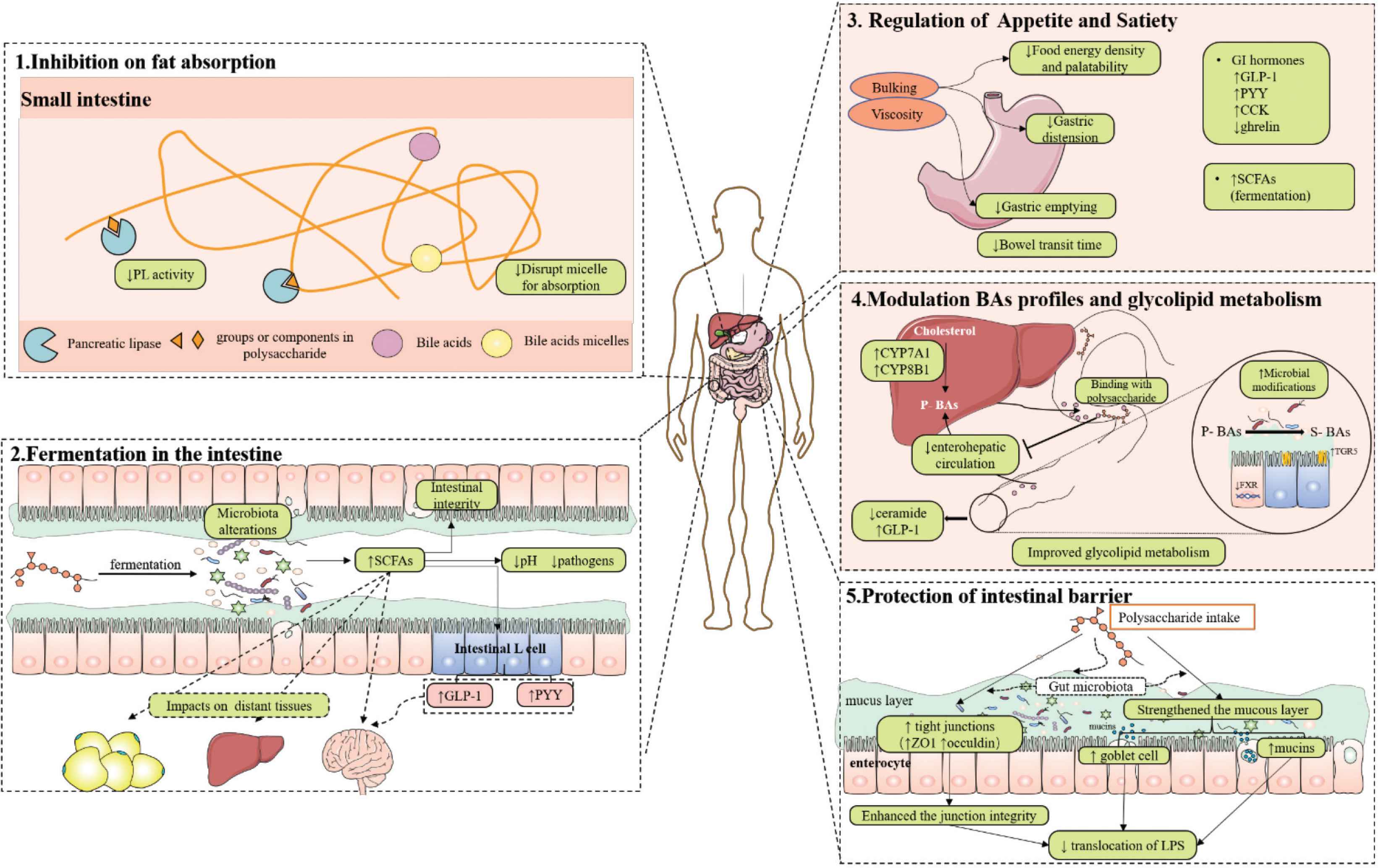
Schematic of mechanisms by which NDPs combat obesity during the gastrointestinal process. (1) NDPs regulate fat absorption via targeting the activity of PL and BAs micelles; (2) NDPs modify gut microbiota and SCFAs through fermentation in the intestine to exert their health-promoting activities; (3) NDPs enhance gastrointestinal satiety by their physical effects and gastrointestinal hormones formation to lower food intake; (4) NDPs regulate the BA profiles and glycolipid metabolism; (5) NDPs protect intestinal barrier and attenuate metabolic endotoxemia. P-BAs: primary BAs; S-BAs: secondary BAs. Images of GI tract and organs (brain, liver, stomach) are referenced from the medical site http://smart.servier.com/ by Servier licensed under a Creative Commons Attribution 3.0 Unported License.
In conclusion, the anti-obesogenic effects of NDPs during their GI digestion and fermentation are significant and worth further discussion. From Tables 2 to 4, anti-obesity effects and mechanisms of NDPs from different sources with structural and chemical composition differences are shown. At present, there is a lack of clear structure–activity relationships between NDPs and their anti-obesity effect, which should be further investigated. Furthermore, it is worth noting that there are still some factors to be considered for the practical application of NDPs in the prevention of obesity. For instance, how to solve the demand for fat-soluble nutrients when fat absorption is inhibited is worth considering. And the methods of NDPs preparation should be simpler, and economic for industrial production. Finally, consumption pattern [134] and dosage also influence the effectiveness of the NDPs. More toxicological studies are needed to find the optimal dose for clinical application. It is anticipated that new products based on NDPs for weight control after these problems are solved.
CONFLICTS OF INTEREST
The authors declare they have no conflicts of interest.
AUTHORS’ CONTRIBUTION
YZ consulted the literature, drafted and revised the manuscript. QX, PC-KC and ZZ reviewed and edited the article. LY contributed in raising the idea, drafting the outline, revising, final approval and funding acquisition.
ACKNOWLEDGMENTS
The work was funded by the
Footnotes
REFERENCES
Cite this article
TY - JOUR AU - Yajun Zhang AU - Qingtong Xie AU - Lijun You AU - Peter Chi-Keung Cheung AU - Zhengang Zhao PY - 2021 DA - 2021/03/18 TI - Behavior of Non-Digestible Polysaccharides in Gastrointestinal Tract: A Mechanistic Review of its Anti-Obesity Effect JO - eFood SP - 59 EP - 72 VL - 2 IS - 2 SN - 2666-3066 UR - https://doi.org/10.2991/efood.k.210310.001 DO - 10.2991/efood.k.210310.001 ID - Zhang2021 ER -